
ウイルス感染と免疫応答【3】血清学的免疫応答(液性免疫応答)
血清学的免疫応答serological immuneresponseは抗体(正確にいえば免疫グロブリンimmunoglobulin)によって媒介されます.
抗体は血液循環に適合した糖タンパクで,標的とする抗原に特異的な分子構造に結合することができます.
抗体の構造
抗体antibodyはY字型構造をしており,先端に抗原と結合する可変部variableregionを持ちます.
抗原は抗体と結合する分子と定義されていますが, より広く,免疫系に認識される分子構造という意味でも用いられます.抗原は通常はタンパクなのですが,糖などの他の構造も抗原になることがあります.
適応免疫応答を刺激する抗原は免疫原とよばれます.
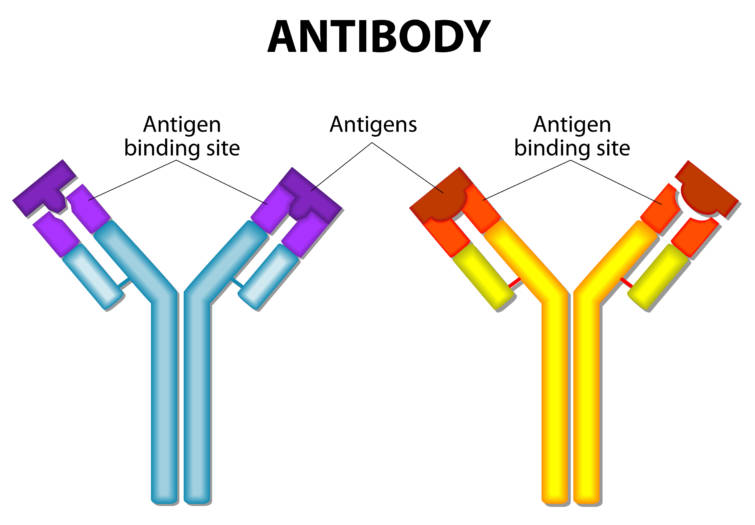
他方,定常部constant regionは免疫工フェクター機能をもたらすための信号を伝える部分です.
抗体は活性化B細胞,特に完全に分化した形質細胞によって産生されます.形質細胞は大きく非常に専門化した細胞です.この細胞が腫瘍化したのが骨髄腫です.
抗体(免疫グロブリン)は2個の軽鎖light chain(短い外側のほう)と2個の菫鎖heavy chainからなる4個のポリペプチド鎖からなります.
抗体は大部分が不変(定常)の構造で,特異的(特異的とは一対一対応のことをいいます)抗原に結合できるよう先端に可変部を有しています.
結合活性は可変部内にある3個の超可変部にあり,軽鎖と重鎖の対になった領域でそれぞれ形成されます.
これらの相補性決定領域complementaly-determining regions(CDR)にはCDR,CDR2,CDR3の3つがあり, これらは集まって特有の抗原結合部位を形成します.CDR3は特に菫要です.
大部分の抗原はタンパクであるが,抗体は脂質や糖などの幅広い分子とも結合することができます.
その結合は非共有結合です.
免疫グロブリンの種類
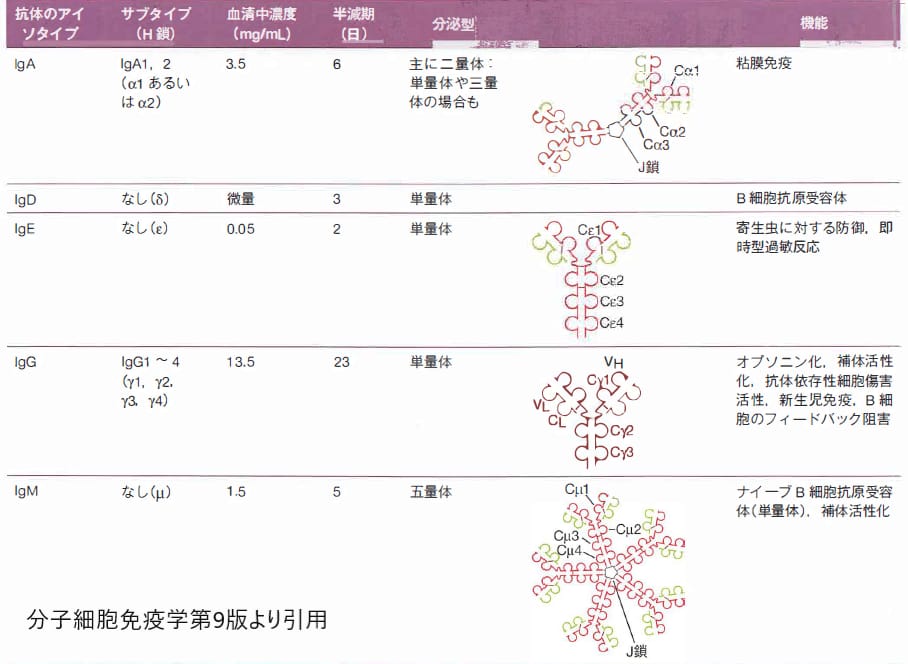
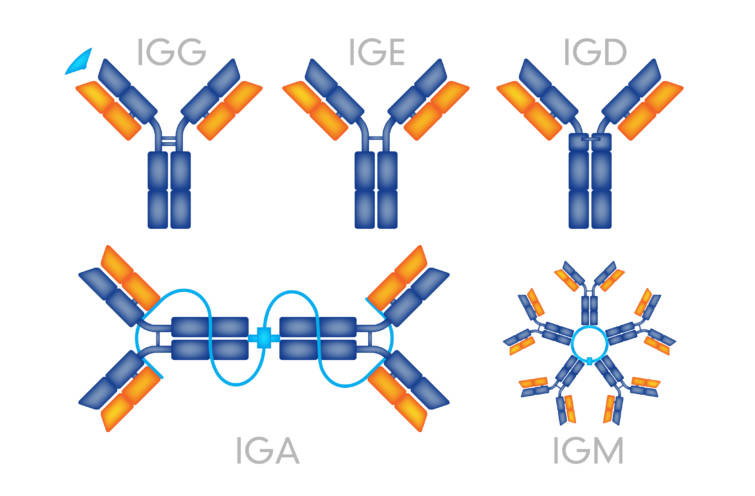
ヒトの免疫グロブリンには5種類の基本的な型があります.
中心となるのは免疫グロブリンG(IgG)です.
親和性が高くて効果的に抗原に結合できる抗体を作るために,多種類の変異型を作ることで適応進化していきます.
免疫グロブリンM(IgM)は5量体で大きいのが特徴です.適応進化を受けないのですが,互いに結合した5個のIgM分子と1個の小さいJ鎖joining chainからなる五量体として血中に存在します.
免疫グロブリンA(IgA)は,血中に存在するが,小胞輸送によって上皮細胞を通過して粘膜表面へ分泌される. このIgAはJ鎖を持つ二量体で粘膜免疫において重要です.
免疫グロブリンD(IgD)は血清中に少室存在するが,その主な機能は,抗原刺激に応答したB細胞の増殖を活性化する受容体の成分の1つとされています.
免疫グロブリンE(IgE)は血清中に非常にわずかしか存在せず,主にアレルギー反応やアナフイラキシー反応に関わっています.
B細胞受容体と増殖反応
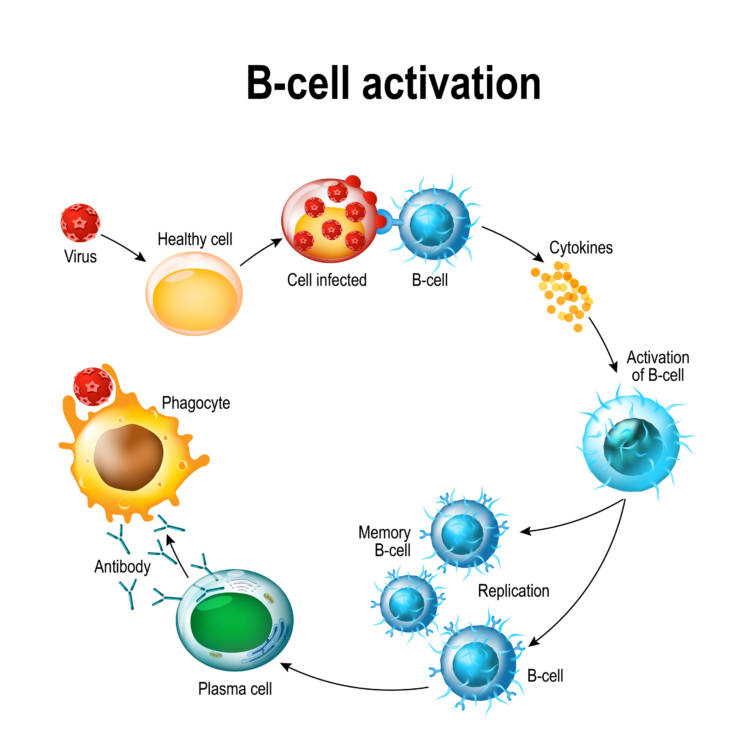
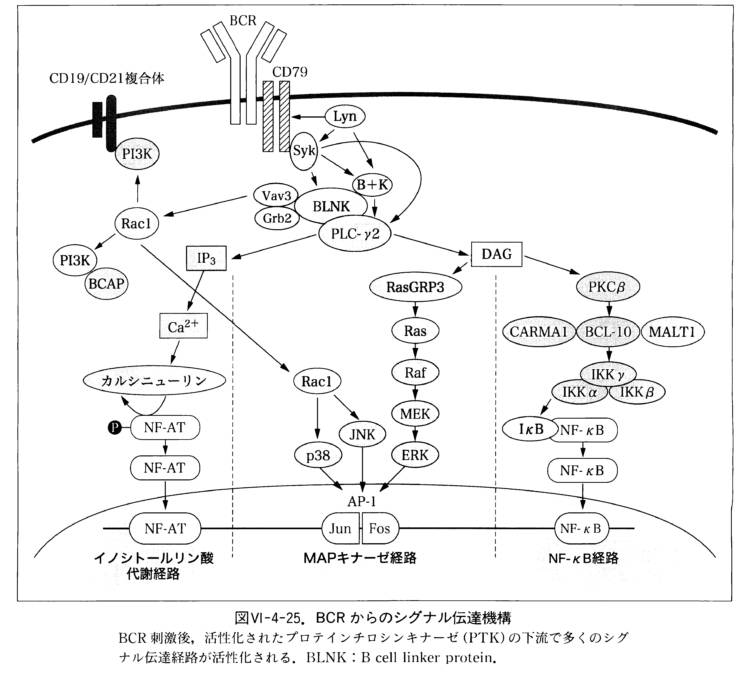
ナイーヴ(未熟)B細胞は,単室体の膜結合型IgDあるいはIgMからなる細胞表面受容体(B細胞受容体B-cellreceptor, BCR)を持ちます.B細胞がその標的抗原と出会うと,抗原がBCRへ結合することにより受容体が架橋され,細胞内シグナル伝達経路が活性化されリン酸化が誘導されることでシグナル伝達がなされていきます.
出典は戸田細菌学第33版です.
強い増殖シグナルを発するには,ヘルパーT細胞によって供給される副刺激がしばしば必要となります.
BCRに結合した抗原は,増殖刺激を与えるだけでなく細胞内にとりこまれてペプチドに分解されて,MHC-Ⅱ分子との複合体となって細胞表面へ輸送されます.
この複合体はヘルパーT細胞上のT細胞受容体T-cell receptor(TCR)によって認識されます.
T細胞が産生するサイトカイン,特にインターロイキンー4 inter-leukin4(IL-4)がさらに増殖するための刺激となります.
B細胞の増殖を誘導するためには副刺激の複雑なシステムが必要であるにもかかわらず,一部の抗原はT細胞の援助なしで同様の作用を生み出すことができるのですが,そういう抗原は細菌の細胞壁に見いだされるような高度な繰り返し構造を持っていることがほとんどです.
抗体の多様性はどうやって作られるのか?
抗原への結合の多様性は抗体が形作られる過程で生じます.
自然免疫系では,あらかじめ決まった範囲の構造のパターン分子しか認識されないのですが,その抗体によって認識される標的抗原は非常に広範囲となっています.
抗体分子の可変部は特異的な遺伝子によるコード(非常に多くのDNAを必要とし,新しい抗原に
対応する柔軟性がない)ではなく,多数の小さな遺伝子の断片によってコードされています.
B細胞の成熟化の間に,複数の可変部断片が選択的に合体することで抗体ができます.
免疫グロブリン鎖をコードする遺伝子をまとめたものです.
| 鎖の型 | 鎖 | 遺伝子がある染色体 | 可変部断片 | 結合部断片 | 多様性断片 | 定常部断片 |
|---|---|---|---|---|---|---|
| 軽鎖 | κ | 2 | 40(重複) | 5 | ― | 1 |
| 軽鎖 | λ | 22 | 30 | 4 | ― | 4 |
| 重鎖 | 全部 | 14 | 40 | 6 | 25 | 2α 1δ 1ε 4γ 1μ |
個々の遺伝子の断片サイズは多様性断片に対する6塩基から低上部消化管内視鏡検査に対する200万塩基まで様々です.
免疫グロブリンの型を決定する定常部は可変域の下流に位置する遺伝子断片の複合セットで酸性されます.結合する定常部はスプライシング(いらないところを切って捨てる)により変化します.
B細胞は骨髄で生まれ,血中や脾臓に移動し,そこで成熟するB細胞はそれぞれ特定の抗原に対する特有の受容体をコードしている自己抗原を認識する細胞は細胞死(アポトーシス)を受けてなくなります.
受容体がその特異的抗原で架橋されるとB細胞は分裂し, 同の受容体を持つ娘細胞クローンをつくる抗原を結合したB細胞のみが活性化されて増殖します.
こうつした方法により可変部に途方もない多様性が生まれることになります.
原則としてはランダムに作られ,それゆえほとんどいかなる抗原にも対応する抗体ができます.
宿主の自己分子を認識する抗体を産牛する細胞は,成熟化の間にクローン選択clonal selectionと呼ばれる過程で排除され, 自己に反応せず,外因刺激にのみ反応する抗体産生クローンのみが残されるのです.
B細胞が抗原と出会って増殖した後でも, さらなる変異が生じ,より変化して親和性の高い抗体が生じます.
免疫グロブリンの可変部の遺伝子に酵素誘導性の点突然変異(体細胞突然変異といいます)が生じることで, B細胞の多様な変化が誘導されます.B細胞のほとんどは,低親和性で抗原に効果的に結合できない抗体を生じる為,高親和性でより効果的に結合するわずかな抗体産生細胞を残して,それ以外の低親和性の細胞は除去されます. これを親和性の成熟affinity maturationといい, これにより効果的な抗体が産生できるようになります.
すごくないですか??
わたしたちの体の中でミクロな戦士たちがこうやって戦ってるんですよ!外敵と.しかもだんだん強くなり,ヨワイ兵士は淘汰されてるんです. 邪魔だよ?どいて って感じで.
もう,マニアックな洋美ちゃんは嬉しくていとしくてB細胞にスリスリしたいくらいです!!
うっとりー(*///∇///*)
血中での抗体の親和性を測定することができるので,低親和性の抗体が存在している場合には,免疫応答が初期の段階にあるということがわかります.
親和性が成熟するにつれて,可変部をコードする遺伝子が,その近くに存在する異なる定常部をコードする遺伝子と融合することにより,産生される免疫グロブリンのクラスが変化していきます.
最初に産生される抗体はIgMで,その後IgGとIgAに変換されるIgGの4種のアイソタイプとIgAの2種のアイソタイプのうち,どれが産生されるかは, この変換反応を制御するヘルパーT細胞とサイトカインの組み合わせによって正確に規定されています.
出典戸田細菌学第33版
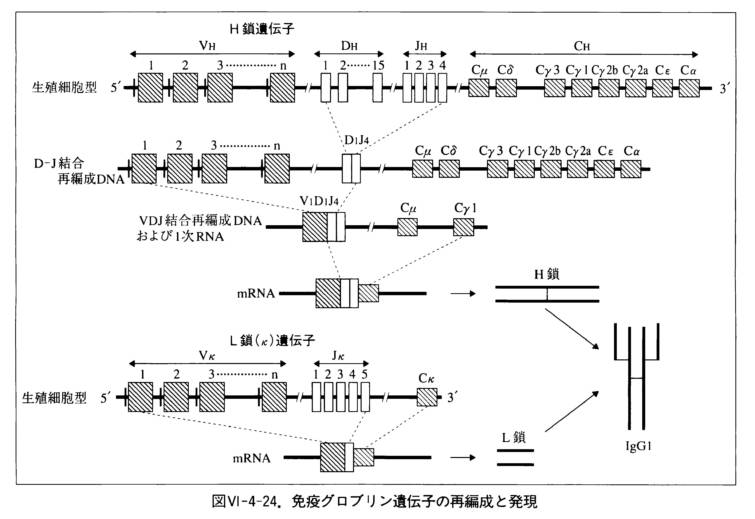
ヒトの抗体分子(とB細胞受容体)は、三つの遺伝子座の遺伝子で暗号化されるC部位とV部位との重鎖(H鎖)・軽鎖(L鎖)で構成されています。
それぞれ、第14染色体の免疫グロブリン重遺伝子座(IGH)、第2染色体の免疫グロブリンκ(カッパ)遺伝子座(IGK)、第22染色体の免疫グロブリンλ(ラムダ)遺伝子座(IGL) に遺伝子断片があります。
つまり、重鎖遺伝子・軽鎖遺伝子は、(V,D,J)三種類の遺伝子断片の組み合わせは、それぞれの遺伝子断片数のかけ算の数だけあり、抗原の多様性に対する抗体の多様性を担保しています。
免疫グロブリン重鎖部位は44個のV 遺伝子断片、D遺伝子断片は27個、J遺伝子断片は6個あります。
そして 軽鎖もまたVおよびJ遺伝子断片から成るが、軽鎖にはD遺伝子断片はありません。
このようにDNA断片をしかるべく選択することを再構成と呼び、この再構成により抗原提示細胞からから遺伝子断片をコピーし、約3000億種類の抗体をつくることができます。
また、T細胞受容体も一つのアルファ鎖と一つのベータ鎖から構成されています。T細胞受容体は免疫グロブリンととてもよく似ていて、V、D、J遺伝子断片をベータ鎖に、V、J遺伝子断片をアルファ鎖にふくみ、それらの遺伝子断片はリンパ球の成長過程で再構成され、そのT細胞に特有の抗原受容体反応を提起します。
これらをIg再構成, TCR再構成といいます。
この変換により抗体の抗原への結合特異性は変化しないが,産生される免疫グロブリンの種類が異なることにより,抗体が免疫グロブリンに結合する結果生じる事象は変化します.
このことは免疫応答の性質と有効性に大きく影響することには注意しないといけません.
特異抗体の存在は患者がウイルスに感染しているかを決定するのに使われることが多いのですが,これもまた出なかったら感染していないということを否定できないことに注意しないといけません.臨床検査の解釈はそう簡単ではないのです.
IgMは感染が最近起きたことを表し,特異的IgGは過去に感染歴があることで起きた免疫応答をあらわします.
免疫グロブリン応答はシステムが非常に多岐にわたるため複雑です.
それぞれの個人の間で, 100万個以上の異なる抗体分子が存在し,それぞれは一連の個人ごとに異なる【感染】経験に応じて生成されますので,異なる抗原に対しては特異的,つまり一対一対応で作られます.
記憶するB細胞
抗原はたくさんあるのですが,同じものに感染しても軽く済みますよね?どういうメカニズムなんでしょうか?
親和性の成熟affinity maturationが終わると,完全に成熟した抗体産生細胞は形質細胞と名前も変わり,骨髄へ移動します.
他のB細胞は記憶細胞memory cellと呼ばれる細胞になり, 同じ抗原がまた入ってくるという再刺激に応答して大量の高親和性の抗体を産生することができます.なので2回目以降は同じ抗原に対する応答は早くなります.
この【特異抗原の再出現】に対する記憶細胞の強力かつ急速かつ大変効果的な免疫応答あは適応免疫の基礎をなすものです.
しかし,新しい抗原が以前免疫応答を生み出した抗原と同じではないのに非常に類似している場合,記憶細胞が見覚えのある抗原に対し最適な効果を持たない抗体を産生することが新しい抗原に対する応答の効率を落としてしまうことがあります.
これを「抗原原罪」”original antigenic sin”といいます.
同様の作用はT細胞でも起こってしまいます.
☛今回のSARS-cov-2ともともと季節的に循環しているコロナウイルスとの間には抗体の交差反応,つまりもともと旧型コロナちゃんに感染した記憶細胞があると新型コロナちゃんに反応してしまうという現象があると報告されています.この辺が理解できるとそのこと自体が決して良いこととは限らないということがわかっていただけまして?免疫って複雑なんですよ.だって女の子なんだもん!(笑)
エピトープ
B細胞は抗原のどこを認識しているのでしょうか?
ウイルスの場合,エンベロープを持つウイルスの糖タンパクのような,ウイルス粒子の外側のタンパクが大体認識される部分です.そうしたタンパクは感染細胞の表面にも存在することが多く,それを認識できます.
免疫応答を惹起する性質を免疫原性といいますが,免疫原性があるタンパクというのは, BCRが結合するための部位をいくつか持ち,こういう抗原として認識される部位を抗原決定墓もしくはエピトープepitopeと呼びます.
エピトープへの免疫応答の順番
さっき,エピトープは複数あるといいましたね?
それらのエピトープヘの応答の順番はウイルス感染に対する免疫応答において非常に菫要となります.
ウイルスはたくさんの抗原を持っているが,免疫応答の仕方は同じではないのです.
免疫を惹起する能力の高いエピトープは特別なタンパクで,特に細胞やウイルスの表面に発現しているタンパクの比較的限られた領域に存在することが多くなっています.
免疫原性の高い領域はが認識されることで高濃度の異なる抗体産生に帰結します.
全てのウイルス抗原の中で, どの抗原が効果的に免疫を誘導するかを予測する方法は今のところほとんどないのですが,個々の抗原への抗体反応を,典型的なウイルス感染患者から採取した血清と比較することで評価します.
こうした点は,ウイルスに対する血清学的免疫検査を設計する時や,ワクチン生産や治療用抗体の作製の際に重要となります.
☛テキトーに作ってもダメなんです,ってことがわかっていただけまして?2位じゃだめなのよ,2位じゃ.
エンベロープを持つウイルスの中和には立体的なウイルスエンベロープが関わっています.
このため,血清学的免疫応答を調べる際にはタンパクが変性して立体構造がかわってしまうような過程をともなう検査はできないため,系が複雑なのだが比較的変性を惹起しにくいものが立体エピトープを調べるのに用いられます.
抗体が結合するとどうなるの?
抗体がウイルスタンパクの特異的に結合すると,ウイルス粒子上であってもウイルスが感染した細胞の表面であっても,大事なウイルス機能が阻害されるとウイルス感染を直接的に阻害(中和 neutralization)する結果をもたらします.
しかし,抗体によるウイルスの特異的中和は基本的には研究室内での状況設定で起こるものです.なぜなら生体内では,結合した抗体のFC領域(定常部)は免疫系の特定の構成要素に対する目印として機能し,そこから先,いろんな反応を引き起こすきっかけになるだけであって,抗体だけでウイルスを中和して終わっているわけではありませんので.
生体内での抗原に対する抗体結合の影響
1.抗体で覆われたウイルスを貪食・消化する食細胞による取り込みの増強(オ
プソニン化opsonization)
2.抗体依存性細胞傷害(ADCC):NK細胞は膜上のFC受容体を介して標的細
胞に近づき,抗体が結合している細胞を殺します.
3.抗体依存性の古典的経路による補体系の活性化:膜の抗体が適切に結合した箇所に致死的な穴をあける,膜侵襲複合体を形成,などの複数の反応が起こります.
4.炎症を惹起し,血管を拡張し貧食細胞を呼び寄せる
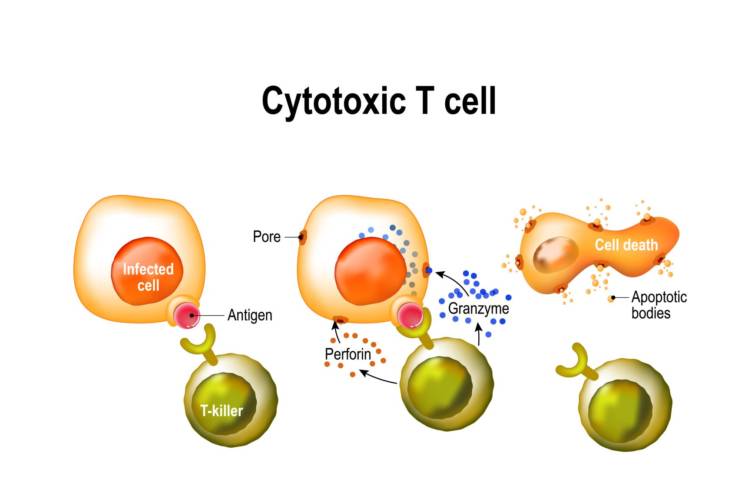
細胞傷害性T細胞は免疫応答を調節し,パーフォリンとザクロ酵素を放出し感染した細胞またはがん細胞を攻撃する.パーフォリンの作用を通じてザクロザイムが細胞質に入り死に至る.
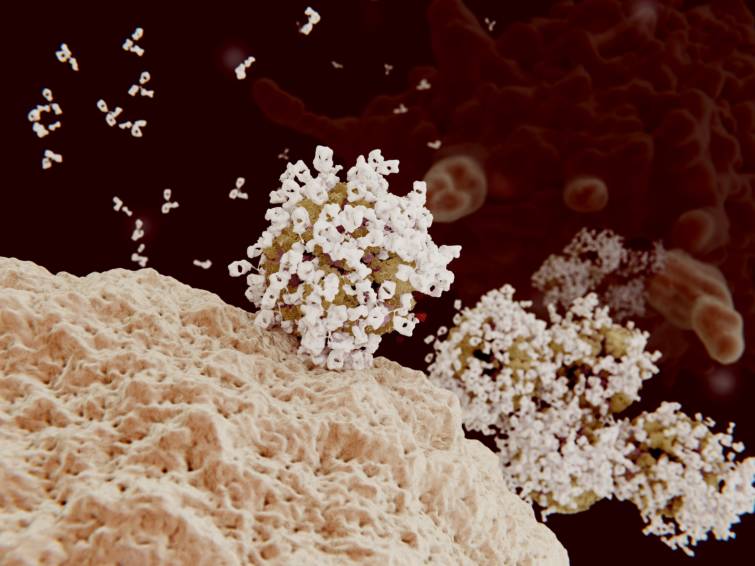
インフルエンザウイルスの抗体オプソニン化の3Dレンダリング.抗体でコーティングされたウイルスは、標的細胞に侵入できずマクロファージ(背景細胞)に飲み込まれて破壊されます.
血清学的応答って何のためにあるの?
血清学的免疫応答(抗体による免疫応答)は,特にポリオウイルスのような細胞外に大量に存在するウイルスの制御に非常に重要となります.
細胞性免疫応答は,感染細胞とその中のウイルスを攻撃できるので,へルペスウイルス科のように細胞内に存在するウイルスに対してより効果的な免疫システムとなります.
ポリオウイルスは,膓管での分泌型IgA産生だけでなく,血中のIgGとIgM産生を含む液性免疫を両方誘導するためウイルスを直接中和できます.
IgG応答は,個人の生涯を通じてずっと持続し,さらなる感染を防ぐ抗体が感染した個人によって産生されるという能動液性免疫に加えて,他人の作った抗体の投与による受動液性免疫もまた可能なのです.
***********
いかがでしたか?
脊椎動物の中でおこっている病原体と免疫担当細胞の戦い.
次はいよいよ細胞性免疫応答です!
まだまだ続くジョ!!
複雑なんだジョ!!


コメント