
ウイルス感染と免疫応答【4】細胞性免疫応答
自然免疫系と抗体が媒介する免疫は,侵入した微生物の表面にある分子を認識することに依存しています.
これに対してT細胞(リンパ球の一種)は,細胞内でタンパクが切断されて生じるペプチド(アミノ酸が2個以上つながったもの)が自己の主要組織適合遺伝子複合体major histocompatibility complex(MHC)分子と結合して細胞表面に提示されたものを認識します.
提示される分子(抗原決定基)の性質により, T細胞への抗原提示の効果が決まります.
抗原提示の主な2つの経路,MHC-IとMHC-Ⅱは異なるエフエクター機構を持ち,異なる応答を誘導します.
1. MHC-I経路
MHC-Iタンパクはほとんどすべての細胞上に存在します.
MHC-I経路による抗原提示は多くの場合,提示細胞内で実際に合成されるタンパクに限定されていて,それゆえMHC-I経路は細胞が感染した時にT細胞応答を発動する経路となっています.
MHC-I分子による抗原提示は,発現しているMHC-I分子と適合するTCRを持ったT細胞のみを活性化する(MHC拘束性MHC restrictionといいます).
結合がうまくいくと,CD8表面マーカータンパクを持つT細胞(CD8+T細胞),主に細胞傷害性T細胞cytotoxic T lymphocytes (CTL)が活性化されます.
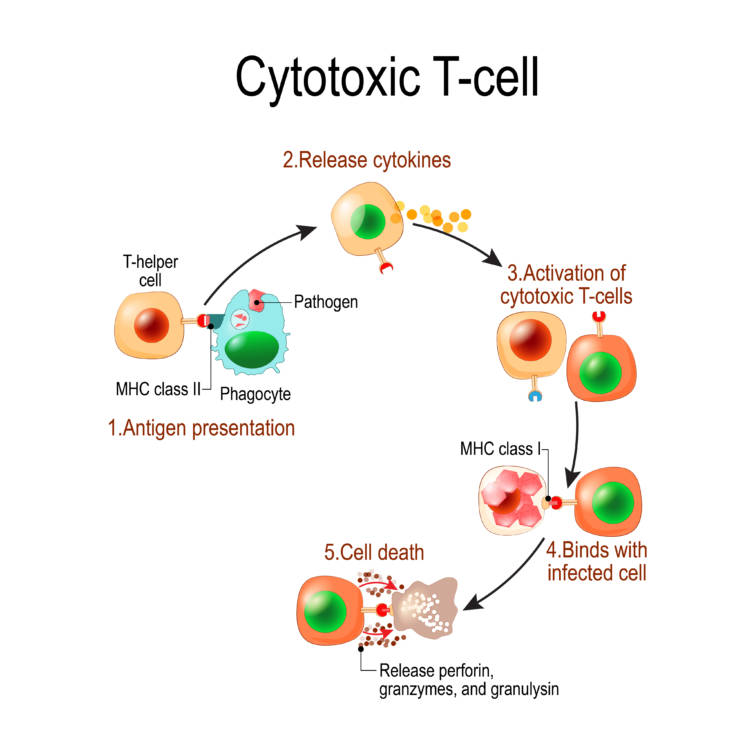
活性化されたT細胞は,サイトカイン産生やパーフォリン(細胞膜に穴をあける物質)の遊離,グランザイム,タンパク分解酵素などによるアポトーシス誘導のような, NK細胞が用いるのと似た方法で抗原提示細胞を殺します.
ほとんどの場合CTLはウイルス感染細胞を殺すことによりウイルスの拡散を防ぎます.
細胞傷害性T細胞は非常に破壊的なため,強く制御されています.
副刺激分子が必要で,副刺激がないと発現する抗原の寛容(免疫系が反応しなくなることをいいます)を導くこと, T細胞応答の働きを修飾するフィードバックシステムの存在などで制御されています.
細胞内の抗原はそこで処理されてMHC-I分子とともに提示され, 抗原提示細胞や同じ抗原を提示している細胞が殺傷されます.この経路を使う細胞は自身を感染細胞と認識し,提示した抗原を標的とする細胞傷害反応を引き起こします.下図はNKcellとなっていますが,CTLと読み替えて結構です.
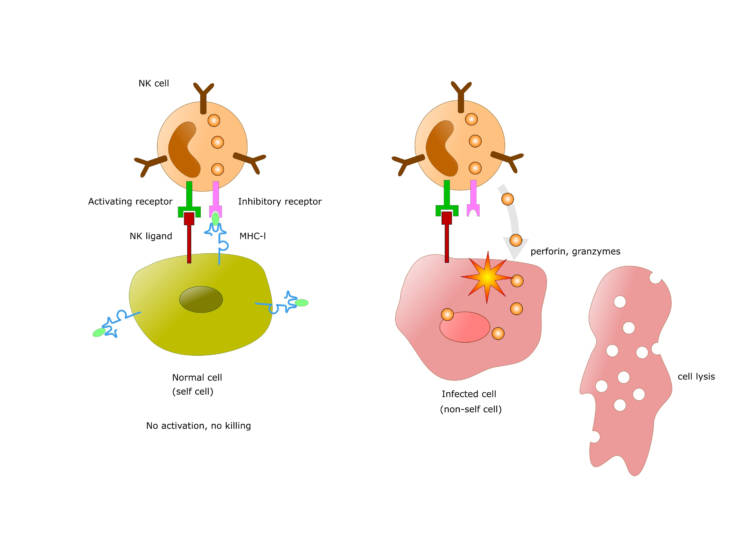
免疫系はこうしてウイルスや病原体が宿主の細胞内に存在しても攻撃することができます.
また,免疫系細胞によって細胞外から取り込まれた抗原は,分解力のあるエンドソームで処理され,MHC-IIと結合して免疫活性化シグナルを伝達します.
T細胞による認識のために提示されうるエピトープは非常に広い範囲に及ぶため,両方のMHCタンパクには多様性が必要となります.
1つの分子構造に特異的に結合する抗体とは異なり,MHCタンパクはペプチド収容溝の基本的性質に適合した一連の異なるペプチドと結合できます.
抗体の場合には結合部位はタンパク, ウイルス,細胞といった立体構造物のいずれにおいてもそれらの表面にあることが普通であるのに対し, T細胞の場合は,タンパク内部のどこからでも,つまり立体構造の内部からでもT細胞に反応するペプチドが作られます. 1つのタンパクに複数のT細胞エピトープが存在し,それは抗体反応を誘導するB細胞工ピトープと大きく異なるのです.B細胞の場合は最終的にそのエピトープに対する抗体を産生するため,同じセルラインの細胞に認識されるエピトープは一つなのです.
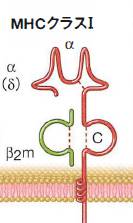
分子細胞免疫学第9版より
MHC-I分子の構造を図示しましたが,深い収容溝binding grooveは特定の構造的な条件に適合した長さ8~10個のアミノ酸からなるペプチドと相互作用できます.
ペプチドは細胞質に存在するタンパク分解酵素複合体のプロテアソームで抗原タンパクが分解されることで生じ,小胞体(ER)を通過してMHC複合体と出会います.
MHC-I経路に入るためには抗原は細胞内で作られなければならないと最近まで考えられていたが,今では,浸透圧ショッ
クや融合性リポソーム,ワクチンアジュバントのなかにも細胞質に入って外来性抗原をMHC-I経路を介して提示するものがあると明らかになってきました.
抗原とMHC-I分子の複合体は細胞表面に提示されます.
2. MHC-II経路
MHC-Ⅱ分子で提示されるペプチドは,MHC-I分子の場合より長く,またバラつきが大きくなっています.
MHC-Ⅱの収容溝がMHC-Iに比べて端が開いているからです.
ペプチドは通常長さ13個以上のアミノ酸からなるが,もっと長くてもよいとされていますが,長いペプチドだとMHC-Ⅱに結合した後,最大でも17個のアミノ酸に切り取られます.
upload.wikimedia.org/wikipedia/commons/thumb/b/bb/MHC_Class_2.svg/225px-MHC_Class_2.svg.png
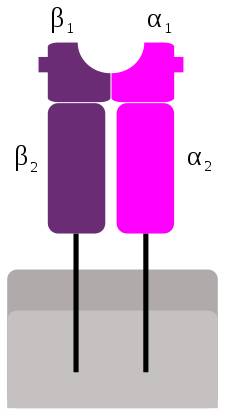
MHC-I経路と異なり, MHC-Ⅱ経路で提示される処理された抗原は,提示細胞内でつくられる必要はなく, また特殊な方法で細胞質に入る必要もありません.むしろ,抗原は特化された細胞で取り込まれ,分解性のエンドソームで分解されたタンパクです.
ペプチド-MHC-Ⅱ複合体は, CD4表面マーカー分子を持つT細胞(CD4+T細胞)にTCR-CD3複合体を介して認識されます.
MHC-Ⅱタンパクは一般に免疫系に密接に関わる限られた抗原提示細胞にのみ発現していますが,皮膚のケラチノサイトのように, ある特殊な環境下に置かれるとMHC-Ⅱを発現することができる細胞もあります.
MHC-Ⅱ経路によって抗原を提示する免疫系の細胞は,異物を童食して他の免疫系細胞に提示します.
それ自身感染細胞ではないので殺されるのは不都合で,CTLを誘導するかわりに,この経路によってヘルパーT細胞helperTcellを活性化します.
抗原刺激に応答してヘルパーT細胞は増殖し,免疫系の抗原提示細胞や他の細胞を活性化するサイトカインを産生します.ヘルパーT細胞とそれが産生するサイトカインは, NK細胞CTL, B細胞などを含む免疫系の多くの細胞成分の活性化に不可欠となっています.ヘルパーT細胞が産生するインターフェロンγ(ガンマ)はMHC-Ⅱを通常発現していない細胞も含め細胞上のMHC-Ⅱの発現を増加させます.
細菌感染した細胞を除去する役割を持つ腫瘍壊死因子(TNF-6)はB細胞に対して抑制的であり,活性化T細胞を殺します.
ヘルパーT細胞によって産生されるサイトカインは,それぞれが複数の機能を持つため,免疫系におけるサイトカインの相互作用は非常に複雑となっています.
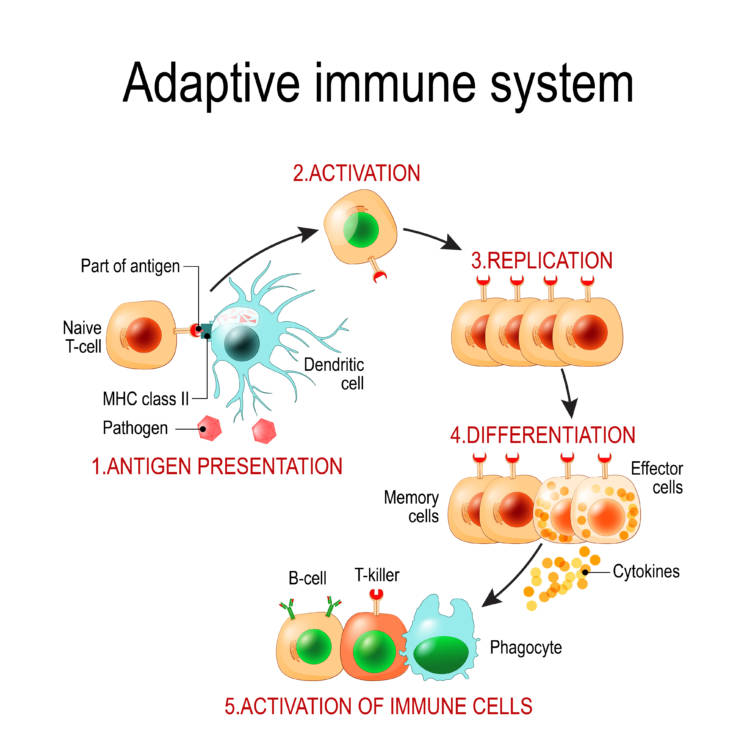
T細胞活性化
T細胞による抗原提示細胞上のペプチド-MHC複合体の認識はT細胞受容体Tcellreceptor(TCR)によって行われます.
TCRは構造が抗体のFa,b領域と似ていて,抗体のように非常に可変性に富む結合領域を持っています. この可変性は複数の遺伝子再編成とTCR分子生成の過程における翻訳機構の組み合わせで生じます.
抗体のように3個の相補性決定領域があるのですが,TCRではこれらのうちの1個のみ(CDR3)が抗原結合に重要な役割を果たします.
TCRはMHC ペプチド複合体に結合してTCRを集合させ,細胞内シグナル伝達系を活性化しますが,この結合のみではT細胞に対して弱い刺激にしかなりません.
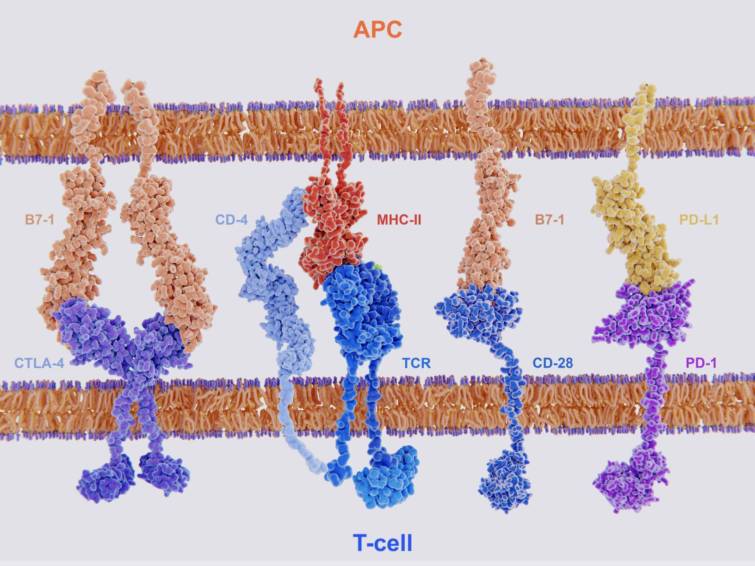
活性化シグナルは, TCR-MHC複合体がT細胞上の他の特定の受容体に結合すると強く増幅されます.
その受容体はMHC-Iの場合はCD8分子,MHC-Ⅱの場合はCD4分子が担っています.
もう1つの重要な副刺激要素がナイーブ(未刺激)T細胞上に存在するCD28が抗原提示細胞の表面に存在するB7タンパクと結合することで,これは, T細胞が増殖するのに必要である免疫系のフィードバック制御をみごとに示すのは, CD28によく似た分
子CTLA-4がこの過程で誘導され, B7とCD28より強く相互作用することです.
CTLA-4とB7との結合は活性化シグナルを遮断し,無規律なT細胞の増殖を防いでいます.
TCR-MHC複合体は直接T細胞にシグナルを伝達しませんが,かわりにCD3複合体CD3 complexと会合している一定の膜タンパクの集まりであるCD3複合体は,細胞内シグナル伝達分子の複雑なカスケードをリン酸化(活性化)し,T細胞へ活性化シグナルを伝達します.
タンパクのなかにははMHC分子による提示されないのにT細胞を直接刺激することができるものがあります.
スーパー抗原(T細胞を非特異的に多数活性化させ、多量のサイトカインを放出させる抗原)はすでに存在するMHC-n-TCR複合体と相互作用することで非常に高度なT細胞応答を誘導し,その結果高濃度のサイトカインが産生され,免疫応答が大きく損傷します.
スーパー抗原は典型的には細菌毒素ですが, ラブドウイルス科の狂犬病ウイルスやへルペスウイルス科のエプスタイン・バーウイルスのようなウイルスにも存在すると想定されますが,それらの役割と性質は細菌のスーパー抗原に比べ不明な点が多くなっています.
ヘルパーT細胞は大きく二つに分かれます.
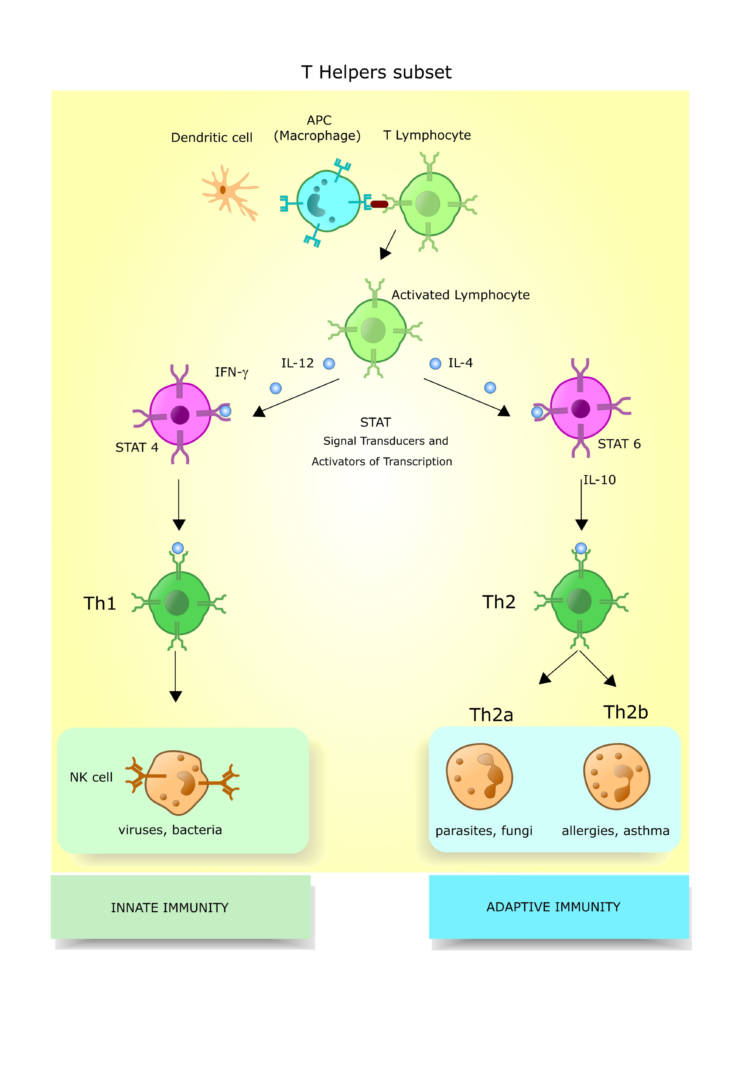
炎症性T細胞(Th1)
細胞傷害と免疫系の炎症応答に関連し,マクロファージの活性化に深く関わります.
Th1細胞はまた, マクロファージを活性化して負食した病原体の破壊を促し,マクロファージの貪食を増強する機能(オプソニン化)を持つ特定のアイソタイプの抗体産生を刺激します.
Th2細胞はB細胞とさまざまな血清学的(抗体)応答を活性化します.

しかし,Th1細胞が特定のタイプの抗体産生を調節しているTh1細胞が活性化されると細胞性,炎症性の応答が優位となり, Th2細胞が活性されると血清学的応答が優位となります.
Th1応答によって産生されるサイトカインとTh2応答によって産生されるサイトカインとは異なっており, これらが応答の性質を決定します.
IL-12, IL-27,TNFα,TNFβ, IFNγの存在はTh1応答.
IL-4、IL-5, IL-6, IL-9, IL-10,IL-13の存在はTh2応答.
***********
いかがでしたか?
細胞たちが病原体と戦うって
感動的ではありませんか?
まさにミクロの戦士.

コメント